| 副目次 |
|
第 2 章 生態系の構造と自己組織化 1 節 生態系の概念と範囲 2 節 物質,エネルギー,用役による相互依存性 3 節 森林生態系の物質・エネルギー流の計測例 4 節 エネルギー流と自己組織化 5 節 最大呼吸仮説 |
| 副目次 |
|
第 2 章 生態系の構造と自己組織化 1 節 生態系の概念と範囲 2 節 物質,エネルギー,用役による相互依存性 3 節 森林生態系の物質・エネルギー流の計測例 4 節 エネルギー流と自己組織化 5 節 最大呼吸仮説 |
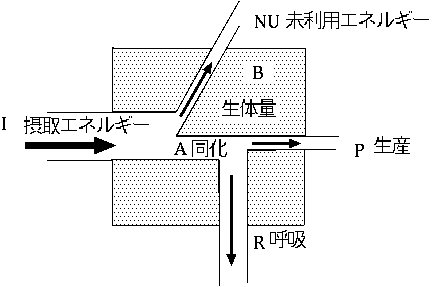
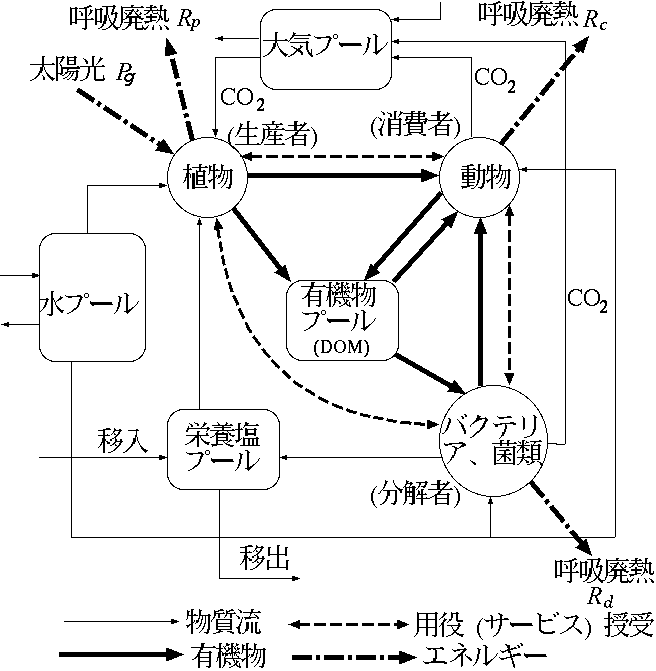
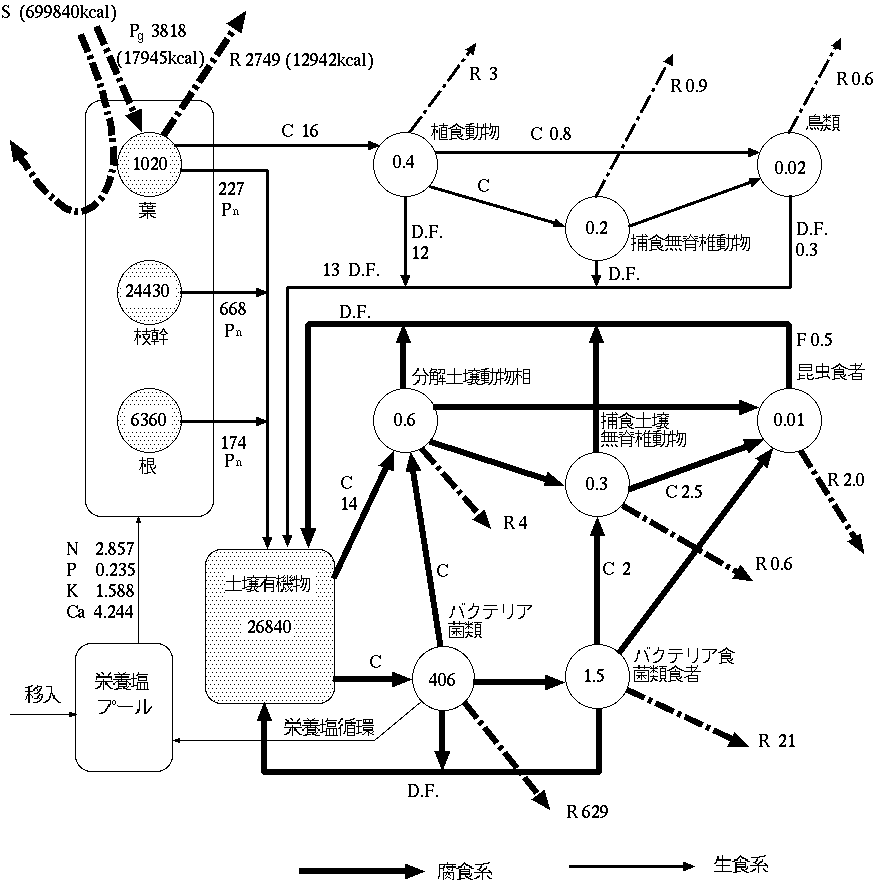
| 林木 | 4.7 |
| 小哺乳動物 | 5.3 |
| 鳥類 | 5.6 |
| 土壌動物 | 5.4 |
| バクテリア・菌類食者 | 5.5 |
| 昆虫食者 | 5.3 |
| N | P | K | Ca | |
| 葉 | 1.723 | 0.142 | 0.466 | 1.374 |
| 枝・幹 | 0.891 | 0.058 | 0.868 | 2.212 |
| 根 | 0.232 | 0.015 | 0.226 | 0.576 |
| 植食者 | 0.011 | 0.020 | 0.028 | 0.082 |
| 計 | 2.857 | 0.235 | 1.588 | 4.244 |
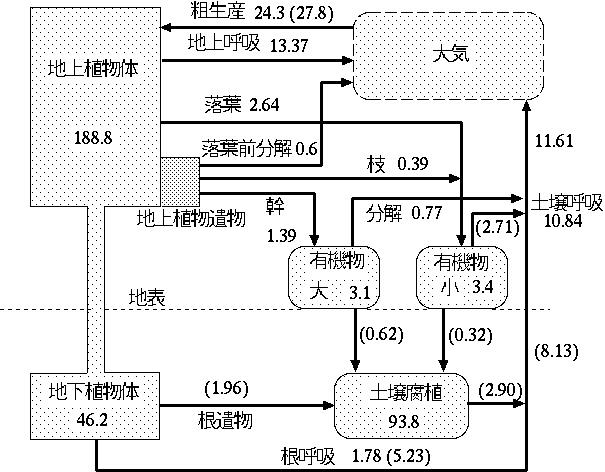

 によって生態学に導入された。(もどる) , p.399。(もどる) 。(もどる) , 東~ , 大串~ , 石川~ など。(もどる) 。(もどる) 。(もどる) を一部改変。(もどる) 。(もどる) を一部改変。(もどる) 。(もどる) , Odum~ 。(もどる) , Jo\llap/rgensen and Mejer~ 。(もどる) 。ただし,ハノンはこの立場をその後も維持しているとは言いがたい。Ulanowicz and Hannon~ 。(もどる) 。この規準によれば,ここでとりあげている原理はすべて, economic-like principle ということになる。(もどる) 。他に Kay~ , Kay and Schneider~ も参照。(もどる) , Kira and Shidei~ , Odum~ , Smith~ 。(もどる) を改変。(もどる) 。(もどる)
によって生態学に導入された。(もどる) , p.399。(もどる) 。(もどる) , 東~ , 大串~ , 石川~ など。(もどる) 。(もどる) 。(もどる) を一部改変。(もどる) 。(もどる) を一部改変。(もどる) 。(もどる) , Odum~ 。(もどる) , Jo\llap/rgensen and Mejer~ 。(もどる) 。ただし,ハノンはこの立場をその後も維持しているとは言いがたい。Ulanowicz and Hannon~ 。(もどる) 。この規準によれば,ここでとりあげている原理はすべて, economic-like principle ということになる。(もどる) 。他に Kay~ , Kay and Schneider~ も参照。(もどる) , Kira and Shidei~ , Odum~ , Smith~ 。(もどる) を改変。(もどる) 。(もどる)