| 副目次 |
|
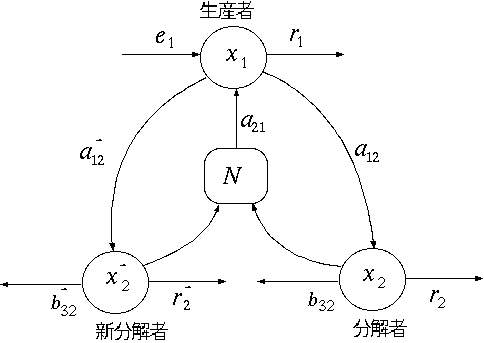
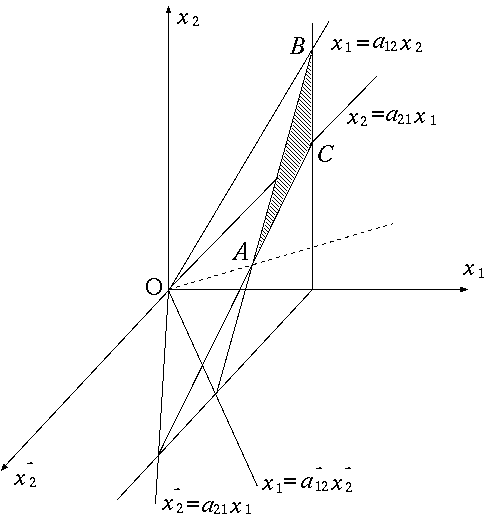
第 3 章 線形モデルによる生態系分析 1 節 線形モデルの可能性と必要性 2 節 生産者−分解者モデル 3 節 分解者間の競争と共存 4 節 最大呼吸構成の安定性 5 節 モデルの一般化と基本定理 |
| 副目次 |
|
第 3 章 線形モデルによる生態系分析 1 節 線形モデルの可能性と必要性 2 節 生産者−分解者モデル 3 節 分解者間の競争と共存 4 節 最大呼吸構成の安定性 5 節 モデルの一般化と基本定理 |
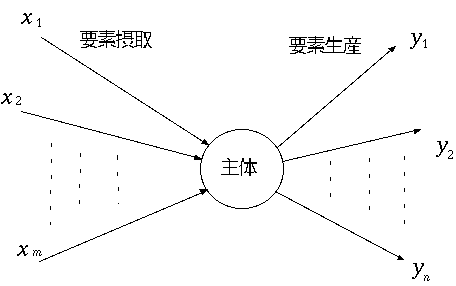
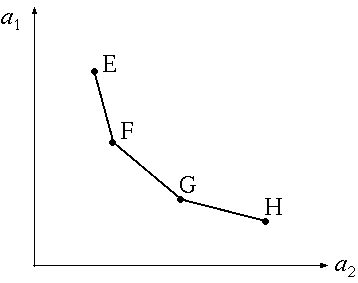
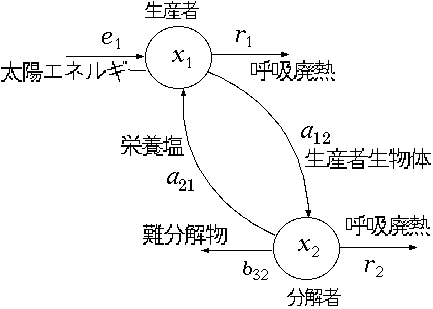
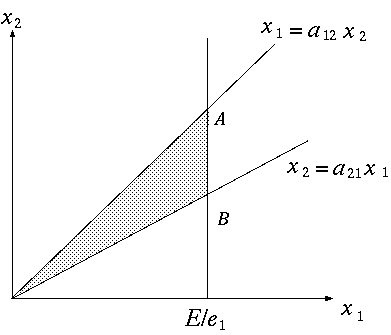

 , p.156。(もどる) 。(もどる) , Finn~ , Hannon~ , Constanza~ , Herendeen~ , Constanza and Herendeen~ , Constanza and Hannon~ , Hannon~ など。(もどる) 。(もどる) , Morishima~ 。(もどる) 。(もどる) 。(もどる) 。(もどる) , p.49。(もどる)
, p.156。(もどる) 。(もどる) , Finn~ , Hannon~ , Constanza~ , Herendeen~ , Constanza and Herendeen~ , Constanza and Hannon~ , Hannon~ など。(もどる) 。(もどる) , Morishima~ 。(もどる) 。(もどる) 。(もどる) 。(もどる) , p.49。(もどる)