| 副目次 |
|
第 5 章 生態系の遷移と動学モデル 1 節 生態遷移の戦略 2 節 モデルの枠組み 3 節 生態系データの導入 4 節 遷移とターンパイク |
| 副目次 |
|
第 5 章 生態系の遷移と動学モデル 1 節 生態遷移の戦略 2 節 モデルの枠組み 3 節 生態系データの導入 4 節 遷移とターンパイク |
| 生態系の属性 | 発展段階 | 成熟段階 | |
| 1. | P/R | 1 より大あるいは小 | 1 に近づく |
| 2. | P/B | 高 | 低 |
| 3. | 食物連鎖 | 直線的 | 網目状 |
| (生食連鎖優先) | (腐食連鎖優先) | ||
| 4. | 全有機物 | 小 | 大 |
| 5. | 種の多様性 | 高 | 低 |
| 6. | 栄養塩の循環 | 開放的 | 閉鎖的 |
| 7. | 細屑物の役割 | 重要でない | 重要である |

| No | 群集構成種 1 〜 19 | P | R | P/R | B | P/B | G |
| 0 | o****************** | 16.67 | 6.67 | 2.500 | 16.67 | 1.000 | |
| 1 | o*oo********o****** | 16.67 | 8.74 | 1.907 | 17.42 | 0.957 | 4.54 |
| 2 | o*oo******o*o*o**** | 1.32 | 3.48 | 0.378 | 2.66 | 0.494 | -84.73 |
| 3 | o*oo******o*o*o**** | 5.97 | 4.18 | 1.428 | 7.11 | 0.839 | 167.39 |
| 4 | o*oo******o*o*o**** | 8.71 | 5.67 | 1.535 | 9.94 | 0.876 | 39.72 |
| 5 | o*oo******o*o*o**** | 8.12 | 5.87 | 1.384 | 9.54 | 0.851 | -4.00 |
| 6 | o*oo*****oo*o*o**** | 8.82 | 8.42 | 1.048 | 11.70 | 0.754 | 22.64 |
| 7 | o*oo******o*o*oo*** | 12.89 | 8.03 | 1.606 | 14.47 | 0.891 | 23.66 |
| 8 | o*oo******o*o*o**** | 8.74 | 6.98 | 1.253 | 10.58 | 0.826 | -26.89 |
| 9 | o*oo******o*o*o**** | 10.91 | 7.72 | 1.413 | 12.82 | 0.851 | 21.14 |
| 10 | o*oo******o*o*o**** | 12.67 | 8.80 | 1.439 | 14.75 | 0.859 | 15.12 |
| 11 | o*oo******o*o*o**** | 13.44 | 9.54 | 1.409 | 15.74 | 0.854 | 6.68 |
| 12 | o*oo******o*o*o**** | 14.68 | 10.40 | 1.411 | 17.20 | 0.854 | 9.26 |
| 13 | o*oo******o*o*o**** | 16.17 | 11.42 | 1.416 | 18.92 | 0.855 | 10.02 |
| 14 | o*oo******o*o*o**** | 17.69 | 12.51 | 1.414 | 20.71 | 0.854 | 9.42 |
| 15 | o*oo******o*o*o**** | 19.36 | 13.69 | 1.414 | 22.66 | 0.854 | 9.43 |
| 16 | o*oo******o*o*o**** | 21.20 | 14.99 | 1.414 | 24.82 | 0.854 | 9.53 |
| 17 | o*oo******o*o*o**** | 23.22 | 16.42 | 1.414 | 27.18 | 0.854 | 9.51 |
| 18 | o*oo******o*o*o**** | 25.42 | 17.98 | 1.414 | 29.76 | 0.854 | 9.50 |
| 19 | o*oo******o*o*o**** | 27.84 | 19.68 | 1.414 | 32.58 | 0.854 | 9.50 |
| .. | ................... | ..... | ..... | ..... | ..... | ..... | .... |
| .. | ................... | ..... | ..... | ..... | ..... | ..... | .... |
| 32 | o*oo******o*o*o**** | 90.59 | 64.05 | 1.414 | 106.04 | 0.854 | 9.50 |
| 33 | o*oo******o*o*o**** | 99.20 | 70.14 | 1.414 | 116.11 | 0.854 | 9.50 |
| 34 | oo*o******o*o*o**** | 108.63 | 77.40 | 1.403 | 126.55 | 0.858 | 8.99 |
| 35 | o*oo*****oo*o*o**** | 119.69 | 110.69 | 1.081 | 156.62 | 0.764 | 23.76 |
| 36 | oo*o*****oo*o*oo*** | 166.67 | 141.24 | 1.180 | 209.51 | 0.796 | 33.77 |
| 37 | o*ooo****oo*o*ooo** | 166.67 | 154.69 | 1.077 | 215.04 | 0.775 | 2.64 |
| 38 | o*ooo*o**oo*o*ooo** | 166.67 | 158.44 | 1.052 | 215.89 | 0.772 | 0.39 |
| 39 | o*ooo*oo*oo*o*ooo*o | 166.67 | 158.89 | 1.049 | 217.62 | 0.766 | 0.80 |
| F | o*ooo*oo*oo*o*ooo*o | 166.67 | 158.40 | 1.052 | 217.24 | 0.767 | -0.17 |
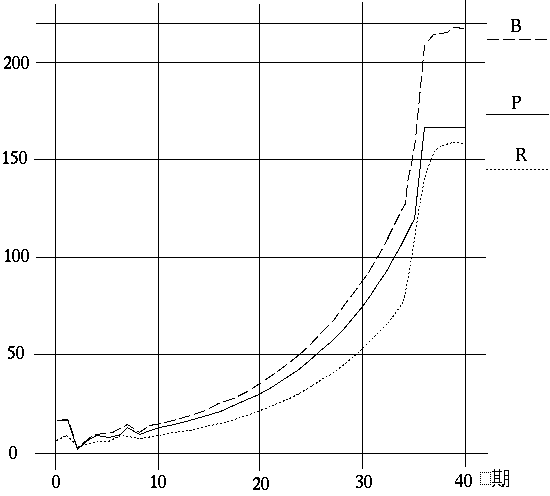
 の指摘にちょうど対応している。すなわち,前者は最終局面が近づくとともに1に近づき,後者は低下している(図~F2)。
の指摘にちょうど対応している。すなわち,前者は最終局面が近づくとともに1に近づき,後者は低下している(図~F2)。 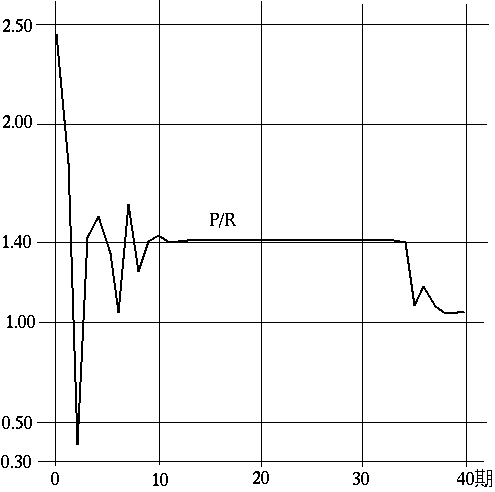
| No | 1 | 3 | 4 | 11 | 13 | 15 |
| 1 | 0.00 | --- | --- | --- | --- | --- |
| 2 | -92.11 | 0.00 | 0.00 | --- | 0.00 | --- |
| 3 | 353.89 | -92.11 | -92.11 | 85.24 | -92.11 | 77.77 |
| 4 | 45.85 | 353.89 | 353.89 | -10.84 | 353.89 | -14.86 |
| 5 | -6.70 | 45.85 | 45.85 | 6.22 | 45.85 | 7.79 |
| 6 | 8.59 | -6.70 | -6.70 | -23.65 | -6.70 | 15.03 |
| 7 | 46.14 | 8.59 | 8.59 | 25.01 | 8.59 | -16.24 |
| 8 | -32.16 | 46.14 | 46.14 | 27.05 | 46.14 | 33.05 |
| 9 | 24.74 | -32.16 | -32.16 | 21.45 | -32.16 | 15.94 |
| 10 | 16.19 | 24.74 | 24.74 | 5.03 | 24.74 | 3.97 |
| 11 | 6.05 | 16.19 | 16.19 | 8.76 | 16.19 | 9.05 |
| 12 | 9.25 | 6.05 | 6.05 | 10.43 | 6.05 | 10.55 |
| 13 | 10.13 | 9.25 | 9.25 | 9.41 | 9.25 | 9.34 |
| 14 | 9.40 | 10.13 | 10.13 | 9.37 | 10.13 | 9.37 |
| 15 | 9.42 | 9.40 | 9.40 | 9.54 | 9.40 | 9.56 |
| 16 | 9.54 | 9.42 | 9.42 | 9.51 | 9.42 | 9.51 |
| 17 | 9.51 | 9.54 | 9.54 | 9.49 | 9.54 | 9.49 |
| 18 | 9.49 | 9.51 | 9.51 | 9.50 | 9.51 | 9.50 |
| 19 | 9.50 | 9.49 | 9.49 | 9.50 | 9.49 | 9.50 |
| .. | .... | .... | .... | .... | .... | .... |
| .. | .... | .... | .... | .... | .... | .... |
| 32 | 9.50 | 9.50 | 9.50 | 9.50 | 9.50 | 9.50 |
| 33 | 9.50 | 9.50 | 9.50 | 9.50 | 9.50 | 9.50 |
| 34 | 9.50 | --- | 9.50 | 9.50 | 9.50 | 9.50 |
| 35 | 10.19 | --- | 9.50 | -25.21 | 9.50 | 5.98 |
| 36 | 39.25 | --- | 10.19 | -38.40 | 10.19 | -11.65 |
| 37 | 0.00 | --- | 39.25 | -40.12 | 39.25 | -42.77 |
| 38 | 0.00 | 0.00 | 0.00 | -13.33 | 0.00 | -27.58 |
| 39 | 0.00 | 0.00 | 0.00 | 0.56 | 0.00 | -86.36 |
| F | 0.00 | 0.00 | 0.00 | -14.13 | 0.00 | 216.84 |
| No | 群集構成種 1 〜 19 | P | R | P/R | B | P/B | G |
| 0 | o*ooo*oo*oo*o*ooo*o | 16.60 | 15.81 | 1.050 | 21.66 | 0.766 | |
| 1 | o*oo****o*o*o*oo*** | 16.67 | 10.97 | 1.519 | 18.91 | 0.881 | -12.67 |
| 2 | o*oo******o*o*o**** | 11.07 | 9.09 | 1.218 | 13.56 | 0.817 | -28.32 |
| 3 | o*oo******o*o*o**** | 14.97 | 10.44 | 1.434 | 17.52 | 0.854 | 29.21 |
| 4 | o*oo******o*o*o**** | 17.12 | 11.89 | 1.439 | 19.92 | 0.859 | 13.69 |
| 5 | o*oo******o*o*o**** | 18.01 | 12.82 | 1.405 | 21.11 | 0.853 | 6.00 |
| 6 | o*oo******o*o*o**** | 19.76 | 13.99 | 1.412 | 23.15 | 0.854 | 9.64 |
| 7 | o*oo******o*o*o**** | 21.77 | 15.37 | 1.416 | 25.47 | 0.855 | 10.05 |
| 8 | o*oo******o*o*o**** | 23.80 | 16.83 | 1.414 | 27.86 | 0.854 | 9.36 |
| 9 | o*oo******o*o*o**** | 26.05 | 18.42 | 1.414 | 30.49 | 0.854 | 9.44 |
| 10 | o*oo******o*o*o**** | 28.53 | 20.17 | 1.414 | 33.39 | 0.854 | 9.54 |
| 11 | o*oo******o*o*o**** | 31.24 | 22.09 | 1.414 | 36.57 | 0.854 | 9.50 |
| 12 | o*oo******o*o*o**** | 34.21 | 24.19 | 1.414 | 40.04 | 0.854 | 9.49 |
| 13 | o*oo******o*o*o**** | 37.46 | 26.49 | 1.414 | 43.84 | 0.854 | 9.50 |
| .. | ................... | ..... | ..... | ..... | ..... | ..... | .... |
| .. | ................... | ..... | ..... | ..... | ..... | ..... | .... |
| 24 | o*oo******o*o*o**** | 101.66 | 71.88 | 1.414 | 118.99 | 0.854 | 9.50 |
| 25 | oo*o*****oo*o*o**** | 111.32 | 85.49 | 1.302 | 133.59 | 0.833 | 12.27 |
| 26 | oo*o*****oo*o*oo*** | 131.22 | 117.93 | 1.113 | 168.63 | 0.778 | 26.23 |
| 27 | oo*oo****oo*o*ooo** | 166.67 | 147.00 | 1.134 | 211.98 | 0.786 | 25.70 |
| 28 | o*oo**oo*oo*o*ooo** | 166.67 | 155.95 | 1.069 | 215.76 | 0.772 | 1.78 |
| 29 | o*ooo*oo*oo*o*oo**o | 166.67 | 160.07 | 1.041 | 218.75 | 0.762 | 1.39 |
| F | o*ooo*oo*oo*o*ooo*o | 166.67 | 158.40 | 1.052 | 217.24 | 0.767 | -0.69 |
| No | 1 | 3 | 4 | 11 | 13 | 15 |
| 1 | 0.40 | -0.40 | -0.50 | 256.69 | -0.39 | 507.39 |
| 2 | -33.56 | 0.40 | 0.50 | 66.08 | 0.40 | 30.27 |
| 3 | 35.16 | -33.56 | -33.56 | 19.46 | -33.56 | 10.59 |
| 4 | 14.36 | 35.16 | 35.16 | 3.68 | 35.16 | 2.67 |
| 5 | 5.24 | 14.36 | 14.36 | 9.37 | 14.36 | 9.81 |
| 6 | 9.71 | 5.24 | 5.24 | 10.53 | 5.24 | 10.61 |
| 7 | 10.17 | 9.71 | 9.71 | 9.30 | 9.71 | 9.21 |
| 8 | 9.32 | 10.17 | 10.17 | 9.38 | 10.17 | 9.39 |
| 9 | 9.43 | 9.32 | 9.32 | 9.56 | 9.32 | 9.57 |
| 10 | 9.55 | 9.43 | 9.43 | 9.51 | 9.43 | 9.50 |
| 11 | 9.50 | 9.55 | 9.55 | 9.49 | 9.55 | 9.49 |
| 12 | 9.49 | 9.50 | 9.50 | 9.50 | 9.50 | 9.50 |
| 13 | 9.50 | 9.49 | 9.49 | 9.50 | 9.49 | 9.50 |
| .. | .... | .... | .... | .... | .... | .... |
| .. | .... | .... | .... | .... | .... | .... |
| 24 | 9.50 | 9.50 | 9.50 | 9.50 | 9.50 | 9.50 |
| 25 | 9.50 | --- | 9.50 | 0.83 | 9.50 | 9.50 |
| 26 | 17.88 | --- | 9.50 | -26.52 | 9.50 | 0.05 |
| 27 | 27.01 | --- | 17.88 | -42.52 | 17.88 | -20.69 |
| 28 | 0.00 | --- | 27.01 | -39.31 | 27.01 | -40.63 |
| 29 | 0.00 | 0.00 | 0.00 | -9.41 | 0.00 | -89.75 |
| F | 0.00 | 0.00 | 0.00 | -5.76 | 0.00 | 238.76 |
, 鷲田~ など。(もどる) 。(もどる) に報告されている。この報告の内容と,本章で提起する最大呼吸仮説による生態遷移の過程の分析との関係についての研究は,今後の課題の一つである。(もどる) , 田川~ など。(もどる) はミクロコズム*における生態遷移の実際の過程を実験した結果として,成熟期における生態系構成種の多様性を指摘している。(もどる) 参照。(もどる)