TOP目次
「環境と社会経済システム」
副目次
0.1 地球的時間のモデル化
0.2 共存共貧のシステム
0.3 システムの冗長性
0.4 自立分散システム
0.5 資源とシステム
補 論 ミクロコズムからの視点
0.1 地球的時間のモデル化
(
副目次へ
)
本書の各章では,今日の自然環境問題と社会のかかわりをいくつかの側面から分析している。そこでは,社会の構造を歴史的に類型化することによって,社会システムという社会のあり方が自然環境問題の解決を困難にしていること,そして日本人も遠い過去に捨ててしまった人格的ネットワークを基礎にした社会のあり方の部分的復権のなかに問題解決の糸口があることなどを指摘してきた。この人格的ネットワークという要素を部分的にでも体現した社会は,小規模な相対的に自立した社会の単位が分権的に社会を構成するという要素をもっていなければならないものである。
本書では,このような視点で,社会の現在ばかりではなく,部分的には社会の過去および未来の描写も試みている。縄文時代
*
から弥生時代
*
にかけての社会システム
*
の形成期についての分析は,確かにそれは発掘された考古学的な事実に立脚しているとはいえ,文字のない時代のことであるから推論が重要な役割を発揮せざるをえなかった。また,自然環境問題の深刻化が引きおこす事態と,それにたいする社会の対応がもたらす社会構造の変化についての議論もまた,それが近い将来のことであるとはいえ,推論が意味をもたざるをえなかった。このように,私たちが自然環境の問題を社会の構造と関連させて考えるときに,なぜ推論という方法が前面に出てくるのか。それは,自然環境問題が人間や社会の存在を非常に長い時間的視野のなかで考えることをよぎなくさせるからである。
あらゆる科学が,人間の存在を相対化する意味をもつことは真実である。どのような科学にも,人間の存在条件や意味を語る側面が存在するのである。自然環境の問題を対象とする科学は,一つは人間と自然のかかわりそのものが人類の誕生以来つづいてきた事実によって,さらに人間が現在自然に与えている影響が深刻かつ長い時間的視野のなかであらわれてくるために,5年や10年という時間のなかで問題を考えることを拒絶するのである。
一方,一人の人間に与えられている時間はきわめて限られている。ほんの数十年の人生の体験によってもたらされる自然と社会にたいする洞察力は,科学や歴史など人類の知的遺産を学ぶことによって研ぎすまされるといえ,求められる時間的視野の長さからいえばはなはだ貧しいといわざるをえない。このようなギャップが今日の環境問題にたいする人々の認識の鈍感さの重要な要因になっていることは否定しがたい。
このような時間的視野の困難を克服するために,関連した歴史や自然科学などの知識をより豊かにすることが大切なことはいうまでもないが,このような時間の流れを実験的モデルによって疑似的に示すことができれば,認識の欠落やギャップを補完するうえで大きな役割を果たすことは確実である。一般に,実験的モデル
*
は複雑な現象や実験が不可能な現象について,問題の基本的な側面をとらえるために採用される,現実に似せながら単純化したものをさす。しかし,時間の流れの悠久さをモデル化することは一見,途方もないもくろみのようであるが,もともと時間の流れは物質の運動の属性のようなものであるから,物的な対象の単純化が時間の流れの単純化も生み出すはずである。
ここでは,このような実験的モデルとして栗原康によって分析されたミクロコズム(微小な宇宙)
*
をとりあげよう(
注1
)。これは,500ccほどのフラスコのなかにつくられた人工生態系
*
であるが,生産者,消費者,分解者といった生態系の基本的な構成要素をもち物質的には閉じた空間のなかで生態遷移
*
をへて極相
*
の定常状態を維持する能力をもっているのである(
注2
)。もちろん,物質的には閉じていてもエネルギー的には外部から光が加えられ廃熱を放散するという点で開放系となっている。この点では,地球と基本的に同じ条件となっている。そして,このミクロコズムのなかで,いま地球的規模で問題になっている持続可能な状態とその条件を実験的に明らかにすることができる。このミクロコズムは外部からの操作によってさまざまな反応をみせる。それによって,システムを構成する主体がつくる構造がどのように定常状態に影響を与えるのかがわかってくるが,その結果は私たちの社会の構造にたいする貴重な視点を供給しているのである。
このミクロコズムは,十数種類の無機塩類
*
と0.05\%のペプトン
*
(タンパク質の分解物)を混ぜた培養液に,ある決まった生物群集を微量移植することによってつくられる。この液に,蛍光灯を12時間照射し12時間暗くするのをくりかえし,温度を24度で維持すると,決まった生物種類による定型的な生態遷移が発生する。まず最初にバクテリア
*
がペプトンを食べて勢いよく繁殖し,しばらくして単細胞動物
*
である原生動物
*
が増加する,バクテリアの減少とともに単細胞で葉緑体
*
をもつクロレラ
*
が増加し,クロレラがピークに達し,原生動物の減少が止まったころにランソウが発生し,最後に多細胞動物
*
であるワムシ
*
が登場する。
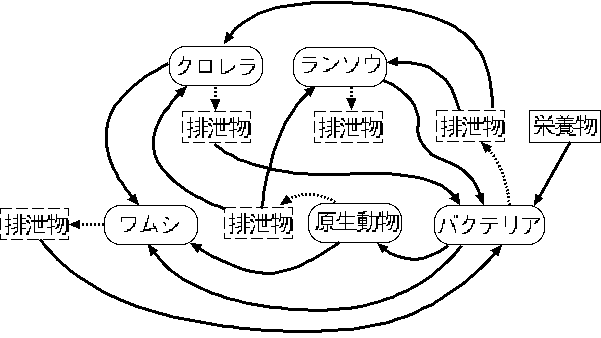
先の生態系
*
のなかでの機能による区分でいえば,クロレラやランソウは光合成
*
が可能であり,生産者に属する。バクテリアや原生動物は有機物を食べたその排泄物の多くが生産者の無機塩類として利用されることから分解者に分類できる。また,ワムシはこの生態系の食物連鎖の最も上位にいる動物であり,消費者に分類してよいだろう。食物連鎖だけに注目したこれらの生物どうしの関係は図~F1 のようになる(
注3
)。図で,点線の矢印は排泄物の生産をあらわし,実線は一方の生物がもう一方の生物または排泄物を食糧にしている関係をあらわしている。
図(F1)
ミクロコズムの食物連鎖
ただしこれ以外にも,これらの生物のあいだには,競争や,排泄物による他の生物の抑制など複雑な関係が存在している。
そして生物群集の移植からおよそ40日で,このような5種類の生物からなる決まった定常状態
*
に到達する。このような定常状態を構成する種は,遷移
*
の途上でさまざまな生物がこのミクロコズム
*
のなかに侵入しているにもかかわらず,変わらないのである。それは,基本的に一定のものとして与えた培養液の栄養の構成にのみ依存する。そして,これらの種のあいだの物質的授受,あるいはその他のすべての関係の結果として,きわめて自立性の高い一つの構造を約40日かけて生成するのである。
すなわち,このミクロコズムは,数百年から数千年の展望でみなければならない地球の閉鎖系内での生物の変遷を,数十日間でみせてくれるのである。もちろん,すべての実験モデルがそうであるように,現実のすべての側面をモデルによってとらえることは不可能である。地球の巨大さと複雑さに比べるとあまりに単純であるこのミクロコズムでとらえることのできる問題はきわめて小さい。しかし,それは物質的に閉じたシステム
*
のなかで,生物が持続的に存在しつづけることが可能であるための基本的な条件を私たちに示している。この重要性は決定的に大きいのである。この実験が明らかにしたもののうち,システムの構造にかかわるものを以下でいくつか取り上げて,自然環境と社会の構造との関係についての私たちの知識にふくらみをもたせることにしよう。
0.2 共存共貧のシステム
(
副目次へ
)
このミクロコズムにかんする実験から引き出された最も重要な教訓は,物質的な閉鎖系
*
が生物にとって持続可能であるためには,共存共栄ではなく,それに属するすべての生物が自己の活力を強く抑制することが必要である,すなわち共存のためには共貧
*
が求められるということである。しかも,そのような共存共貧
*
の状態において,ミクロコズムは外界からの攪乱にたいして最も強い安定性を示すのである。
この教訓がミクロコズムからどのように引き出されたかにさきだって,これが私たちに教えていることを敷衍しておいた方がよいだろう。すでに述べたように地球もまた物質的に閉鎖系
*
である。したがって,そのなかである特定の生物種が繁栄し増加しつづけることはできない。具体的には,人類もまたその活力を最大限に発揮した状態では地球という閉鎖系は崩壊せざるをえないこと,それを回避するためには,人類がその活力をなんらかのかたちで抑制しなければならないことをこのミクロコズムは教えているのである。
私は本書の本文中で,自然環境とのバランスを回復するためには,自然環境の物質的制約を経済が受け入れることが不可欠であり,それは経済の活力そして社会全体の活力を低下させることになると指摘している。そして,このような活力の低下のなかでいかに経済と社会を全体的な崩壊から回避するのかが私たちが直面している最も重要な課題であることを明らかにしている。この私たちの社会に要求されている活力の抑制はミクロコズムのなかで生物種の個体群に要求されている活力の抑制と本質的には同じものである。私たちの社会は,環境の長期的な制約を無視し,直面する制約だけを意識している限りさらに一定期間物質的繁栄とそれを可能にする経済的な活力の増進を実現することが可能だろう。しかし,将来的に社会の持続可能性を失わないためにはこの時期に活力の自己抑制が必要になっているのである。
ミクロコズムのおける定常状態における共貧
*
とは次のような意味である。
先に述べたように,ミクロコズムは遷移
*
の開始した後,基本的な構成種が次々にその個体群の規模を増加させる。生産者による1日あたりの有機物の生産の合計を総生産量と呼ぶことにしよう。このミクロコズムのなかで有機物を無機物から合成することができるのは生産者だけであるからこの総生産はこのシステム全体としての生産量である。消費者や分解者は,この生産物から生産者自身が自己の生体の維持のためにその一部をエネルギーとして用いた,すなわち呼吸廃熱化
*
した残りを利用するだけである。消費者や分解者に利用される総生産も,その一部は彼らの生活の維持のために呼吸廃熱化され,残りが彼ら自身の生物体の増加のために用いられる。したがって,総生産はミクロコズムの生物によって生活のために呼吸廃熱化される部分と,生物体を増加させるために用いられる部分(この部分を純生産物と呼ぶ)に分けられる(
注4
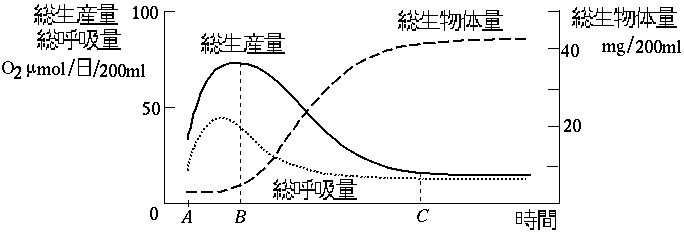
)。このような,呼吸廃熱化と総生産がこのミクロコズムの生態遷移のなかでどのようにしてあらわれたかを,図~F2 に示している(
注5
)。\setcounter{appfigure}{2}
図(F2)
遷移における総生産,総呼吸,総生体量
図において,総生産量と総呼吸量(群集総呼吸)
*
の差は純生産
*
をあらわしている。総生産がピークになるまでこの純生産量は少しずつ増え,それにつれて生態系全体の生物体量も徐々に増加している。図で第一に注目すべき点は,総生産と総呼吸のピーク(B点)が遷移の初期にあらわれていることである。そして,第二に総生産と総呼吸は成熟期(C点より後)にはほとんど等しくなり,システムのなかへの生体の蓄積がおこなわれなくなり,クロレラやランソウによって生産された有機物に体化したエネルギーは,これらの植物自身とそれを栄養源にしている消費者と分解者によって利用しつくされてしまうということである。
では,遷移の初期になぜ成熟期と比べる高い総生産と総呼吸のピークが存在するのだろうか。このピークは,この時期にシステムが全体として生産においてもその利用においても高い活力を有していることを示している。このおそらく最も重要な理由は,500ccという小ささではあっても,そのなかにさまざまな利用可能な初期資源が存在してきたことである。 遷移の出発点にペプトンという資源が与えられて,これが初期のバクテリアの栄養源となり急速な生体の増加を生み出し,バクテリアはさらに原生動物の食糧となり,初期賦存の無機塩類とバクテリアの分解物がその後のクロレラやランソウの栄養源となりえたのである。さらに,このフラスコの空間という資源にも余裕があり,他の生物種との競争やバランスの制約をあまり受けることなく生体量を増やすことができたのである。
しかしこのような初期の余裕は徐々に失われる。そして,それとともにミクロコズムは生物種間の適切なバランスのもとでのみ維持することができるようになる。それは,初期の資源のような早晩枯渇する資源ではなく,すべての生物が他の生物のおこなった生産や分解の結果としての再生資源を利用するばかりでなく,みずからも必ずなんらかのかたちでこの物質循環に貢献しなければならなくなっていくことを意味しているのである。このようなことが要求されるのは,このミクロコズムが物質的に閉じたシステムとなっているからである。そして,このような生物種間のバランスが強制されることによって生物種の全体はもちろん特定の生物種においても活力の増加を持続することは不可能になり,自由な資源の存在する状況のもとで高い活力を発揮した生物種もより低い水準にそれを維持しなければならなくなるのである(
注6
)。
0.3 システムの冗長性
(
副目次へ
)
ミクロコズムがもたらしたシステムについてのもう一つの重要な視点は,システムの冗長性(redundancy)
*
がシステムの安定性と強靭性をもたらすということである。この冗長性には二つの意味がある。一つは,貯蔵資源を保持することによる冗長性である。もう一つは,構造や主体の多様化による冗長性である。
この冗長性はミクロコズムのなかで次のようにあらわれている。まず貯蔵資源の問題であるが,栗原は宇宙基地システム
*
という仮想的なシステムとミクロコズムを比較する。宇宙基地システムのなかでは,人間を生存させるために必要な植物による生産から人間も含めた動物の排泄物まで,物質フロー
*
が厳密に管理されなければならないことを明らかにする。そして,このようなフローのバランスがなんらかの障害にあうと,システム全体が一挙に崩壊する危険性をはらんでいる緊張したシステムであると指摘する(
注7
)。このようなシステムと比べて,ミクロコズムが安定な持続性をもちえているのはシステムの内部に貯蔵系
*
が存在するためであることを解明している。
具体的には,まずミクロコズムにおいては成熟するにしたがって,フラスコの底に生物の死骸が固形物としてたまってくる。フラスコの生物が減少すると,バクテリアによってこの固形物が消費されミクロコズムの活力を増加させ,生物が増加すると,死骸が増加しここに貯蔵されるというかたちで,システムのフローのバランスを調整しているのである。それはちょうど電気回路におけるコンデンサーのような役割を果たしていることになる。
さらに,このミクロコズムは外界の大気から完全に遮断されても光エネルギーだけでシステムを生きたまま維持できるのであるが,遮断された内部の気体もまた貯蔵系として機能しているのである。この気体という貯蔵系の存在もまたシステムの持続能力に大きな影響を与えている。この内部気体の量があまりに小さすぎるとミクロコズムは崩壊してしまうのである。たとえば,フラスコの内部に気体をおかない状態にしてしまえば,いくら生物の物質バランスが維持可能であるといっても,ミクロコズムはやがて崩壊してしまう。すなわち,大気もまた貯蔵系
*
として機能しているというわけである。
もう一つの,主体と構造の多様性にかんする冗長性
*
についてもみておこう。すでに指摘したようにミクロコズムは成熟するにしたがって外部からの攪乱にたいするより高い安定性
*
を備えるようになる。これは,一つには成熟するにしたがって,ミクロコズムを構成する主体の種類が増加することと関連している。種の増加がミクロコズムの安定性にどのように影響を与えるかは必ずしも明確ではないが,最も考えられることは,生態系を維持するために必須の機能が複数の生物種に担われるために,一つの種の機能における障害が他の種によって代替可能になる点である。たとえば,ミクロコズムにおける最も基本的なバランスは,生産と分解のあいだのバランスである。光エネルギーによって有機物を合成できる生産者は成熟期において複数の種から構成されるようになり,分解の機能を果たす消費者や分解者の種類も成熟期では最も多くなり,それぞれが分解の水準やその他の機能で多様な役割を果たすようになるのである。
ミクロコズムの実験できわめて興味深い点は,これらの構成種の多様性
*
がミクロコズムの構造の多様性
*
と密接に関連していることである。構造の多様性とは次のようなものである。ミクロコズムは,初期のバクテリアの個体群の急速な拡大の時期をすぎると,単純にフラスコ内にそれぞれの生物種が一様に分布することはなくなり,たとえば,培養液の表面と中層そして底の部分などに生息場所の分化
*
をおこない,さらには,遷移の段階に応じて生物種のいくつかが分化した集落のようなまとまりを形成し,フラスコ内の空間的構造を一層複雑にしていく。そして,このような構造が生物種の多様性の出現とちょうど対応しているのである。そして,このような構造の多様性が放射線の照射などの攪乱から構造のなかに展開している生物種を守ることによって,強い復元力
*
を示すことができると考えられている。
このような,ミクロコズムにおける冗長性
*
の機能は,私たちの社会をみるうえでも有効な視点となっている。それは,たとえば経済活動における在庫投資などの機能と比較することもできるが,ここでは,社会システム化したことによって自然との関係において社会がある種の冗長性を失ってしまったという問題を取り上げることにしよう。これまでの章で述べてきたように,社会システム化の初期の決定的な要因は灌漑農耕の開始である。日本において灌漑農耕
*
の開始は主食を農耕によってえる社会の開始だった。すなわち,それ以前も農耕という生業それ自体は存在したであろうが,農耕によって食糧の主要な部分を調達するようになったのは,この灌漑農耕の開始によってである。そして,このような農耕の最も重要な特徴の一つは,自然との関係をいちじるしく単純化していることである。
この単純化は,一面では人間が食糧として依存する種の単純化である。日本の場合は水稲という植物種への依存となった。またもう一面では,それによって自然との関係における構造も単純化した。つまり,灌漑農耕以前は自然との多様な関係を社会がつくり上げていたのである。自然の多様な生産をさまざまな時期に利用することによって,自然の変調からくる危険もまた分散させることができた。つまり,自然との多様なチャンネルを確保しながら人間の生活が営まれていたのである。これにたいして,灌漑農耕は自然とのチャンネルも技術も構造も単純化した。それによって自然とのあいだに冗長性が失われ,自然との緊張した関係が避けられなくなってしまったのである。それは,社会システムの形成にも反映し,個人の自由が社会の全体性の管理下におかれざるをえなくなり,そして人と人との関係もまた,緊張した関係となってしまったのである。
0.4 自立分散システム
(
副目次へ
)
ミクロコズムが教える,安定したシステムにかんするもう一つの条件はシステムが自立した分散構造
*
をもつということである。
ミクロコズムは遷移が成熟期にはいると構造が多様化し集落を形成することはすでに述べた。この集落は500ccのフラスコのなかで2\sim3ミリの小さいもので,底にほぼ等しい距離でパッチ状に分布するのである。そして重要なことは,この小集落
*
にはミクロコズム全体の成熟期を構成するすべての種が含まれ,そこで自立的な物質循環がおこなえるようになっているのである。
さらにこの集落の機能を確認するために,他の条件を同じにしながら一方のミクロコズムは定期的に緩く攪拌しながら遷移をおこなわせ,もう一方は攪拌せずに遷移をおこなわせるという実験をおこなっている。後者は,当然,分散した小集落を形成するが前者は同じように成熟した極相
*
に向かうものの攪拌のために集落は構成しない。そして,両者の成熟期の状態を比較すると攪拌のために小集落を形成できなかったミクロコズムでは小集落を形成したものに比較して生物種が一つ少なくなり,またそれぞれの生物種の個対数も10分の1ほどになってしまっていることが確認されたのである。
この実験が意味していることは,物質的に閉じた空間という厳しい条件のなかにある生物システムにおいては,環境が提供する条件を最も効率的に利用するためにある一定の規模の相対的に自立できる小さな単位が分散するかたちで存在することが望ましいことを示している。そして,そのような自立分散型のシステムになることによってより多様な生存条件を確保することができ,したがってまた先に述べたように外部からの攪乱にたいしてもシステム全体の安定性
*
を確保することができるのである。
このようなシステムに自立分散的な構造をはめ込むことは,第5章でくわしく分析したように日本の縄文以前の社会においておこなわれていたことである。そしてまた,環境制約を受け止めることのできる社会をつくり出すために,私たちの社会がとり入れるべき要素であることも指摘した。このような構造は,自然環境と調和的であることを強制される持続的なシステムにおいては不可避的に実現しなければならないものであることがこのミクロコズムによってもまた確認できたのである。
0.5 資源とシステム
(
副目次へ
)
このミクロコズムが提供した視点として最後に取り上げるのは,資源の存在が生物システムに与える影響である。これは,私たちの社会が石油などの化石資源という豊富な資源に支えられていることの結果を示唆するものとして興味深い。
まず,実験そのものを紹介しよう。一つは,成熟期にあるミクロコズムに自然の水域にある量と比べるといちじるしく多いリン酸塩を入れる実験である。リン酸塩はクロレラやランソウの栄養源であり,とくにこれを加えることによってクロレラはそれ以前の50倍もの量になった。このクロレラの排泄物の増大によって,それを食糧とするバクテリアもかなり増加するが,原生動物やワムシのような動物は姿を消してしまう。
もう一つは,バクテリアの栄養源であるペプトンを成熟期にあるミクロコズムに大量に投入する実験である。その結果として,予想されるようにバクテリアは急速に増加する。しかしそれにともないクロレラやランソウは急速に減少する。本来,バクテリアの排泄物はクロレラやランソウの栄養源であるが,それが過剰になってこれらの植物に毒素として作用してしまった結果である。さらに,原生動物やワムシのような動物も姿を消してしまう。バクテリアはこれらの動物の餌になっていたのだが,これもバクテリアの排泄物の過剰が動物に毒として働き殺してしまったのである。
また,このミクロコズムの最上位捕食者であるワムシを成熟期のミクロコズムに大量に投入すると,そのワムシがクロレラを食べつくしてしまうことによって原生動物も消え,またワムシ自身も消えてしまう。
逆に,初期の培養液に加えていたペプトンの量を5分の1にしてミクロコズムの遷移をおこす実験もおこなっている。すなわち,通常よりも資源の少ない状態でミクロコズムを培養したのである。すると一つは,という種もまた成熟期の構成種となりシステムを構成する主体の多様性が増大する。さらに,成熟期までの期間がそれまでのものよりも半分の日時になってしまうという,ミクロコズムの全体的な効率が増大するという結果がえられている。
すなわち,これらのミクロコズムにかんする一連の実験が示していることは,資源が多いことが逆にシステムを攪乱し,また崩壊させてしまうということである。
このことが,私たちの社会にかんして意味していることは直接的なものである。私たちの社会では自然の豊かさと社会の豊かさが連動しない。この日本をみても,明らかに日本の自然は現在のすべての日本人を養う力をもっていない。自然が貧しかろうと豊かであろうと無関係に経済の豊かさを享受しているのがいまの日本の姿である。このようなことが可能になっているのは,日本人の勤勉さもあるかもしれないが,それよりもひたすら石油という優れた資源のおかげである。自然の資源が貧しくても,あるいは存在してもそれを使わずに経済的に豊かであるのは,石油を利用して大量の工業生産物を生み出すことができるためである。
もちろん,これは日本だけの姿ではなく,現在の世界を構成する工業社会
*
に共通した特徴である。問題は,単に,この石油という資源があとどれだけ使うことができるかということではなくなっている。石油という資源を大量に使うような経済のあり方が自然環境を貧困化させいたるところで崩壊させているのである。これは,大量の資源を与えられたミクロコズムが自己を持続的に存在させる条件を喪失してしまうことと本質的に同じ現象である。資源が存在し,利用可能だから利用しつくすまで利用するというのではなく,資源を地球のもたらしている貯蔵系
*
として理解し,自然の不規則な変動にたいするバッファ(緩衝装置)
*
として利用する,そして資源利用をその点にまで抑制し物質循環に支えられた定常的な経済の方向を目指すことがいま求められているのである。
脚注
(1)以下の議論は基本的に栗原~
をもとにしている。関連する他の論文としては,栗原~
,Kurihara~
など。(
もどる
)
(2)第1章,第4節の1.参照。(
もどる
)
(3)栗原~
,p.60の図を改変。(
もどる
)
(4)生物体の増加は,生きた生物としてばかりではなく,死骸や排泄物の未分解部分としてもあらわれる。(
もどる
)
(5)栗原~
,p.23,図7より。(
もどる
)
(6)「有限の空間ととぼしい資源のなかで安定と共存をはかろうとすれば,個々の生物は,数と活力の低下という代償を支払わなければならない」,栗原~
,p.153。 (
もどる
)
(7)「すべての生物は全体に寄与する「部品」と化していることが明らかである。行動の自由はうばわれ,``人口''は統制され,食物の選択も摂取量も制限され,絶えず外の生物の顔色をうかがいながら,戦々恐々生きつづけなければならず,しかも勝手に死を選ぶことすら禁じられている」,栗原~
,p.141。(
もどる
)
副目次へ
 をもとにしている。関連する他の論文としては,栗原~ ,Kurihara~ など。(もどる) ,p.60の図を改変。(もどる) ,p.23,図7より。(もどる) ,p.153。 (もどる) ,p.141。(もどる)
をもとにしている。関連する他の論文としては,栗原~ ,Kurihara~ など。(もどる) ,p.60の図を改変。(もどる) ,p.23,図7より。(もどる) ,p.153。 (もどる) ,p.141。(もどる)